Cell Biology: Definition of Cell, Types, Structure and Functions
At some point or the other, we’ve all marveled at how diverse the living world around us is. Detailed scientific studies of these organisms only emphasized their diversity even more. However, there also exists unity underlying this diversity – and the theory that emphasizes this unity is the cell theory.
The cell serves as the fundamental structural and functional unit of all living organisms. It is the cell that is a universally common feature in every living organism that ever existed, exists today, and will ever exist in the future. The cellular organization gives rise to a plethora of physiological, biochemical, and behavioral processes that lead to the genesis of life itself.
In order to study these processes in molecular terms, scientists have taken a physio-chemical approach and made use of cell-free systems. They further established this approach by analyzing the elements and compounds present in living tissues. Not only does this method explain the molecular basis of physiological processes, but it also helps us understand abnormal processes that take place during diseases.
The aforementioned method of using the concepts and techniques of physics and chemistry to understand biology is known as reductionist biology. It is a rapidly growing field that is helping scientists unravel the mysteries of the living world faster and clearer than ever. Let us now proceed to understand what exactly a cell in biology is, and what it holds a position of such great significance in the field.
Table of Contents
Cell: The Unit of Life
To put it simply, the cell is what differentiates a living organism from a non-living object. An entity can be considered living if it possesses the basic unit of life – the cell. All living organisms you see around you (and even those you cannot see by the naked eye) are composed of cells. Some of them consist of a single cell and are known as unicellular organisms. Others are composed of numerous cells and are known as multicellular organisms.
However, it is worth mentioning that unicellular organisms are perfectly capable of both independent existences and performing all the essential functions of life. In other words, the capability to live independently does not exist for anything less than the complete structure of a cell.
The cell was first discovered in 1665 by Robert Hooke, an English scientist who first observed a slice of bottle cork under a primitive microscope. Although he only observed the dead cell walls, in this case, he laid down the foundations of cell biology. Rightfully, he is now known as the father of cytology.
Anton von Leeuwenhoek was the first scientist to observe and describe live cells such as bacteria, red blood cells, protozoa, sperms, and so on. Later, Robert Brown discovered the nucleus of the cell. The invention of the electron microscope by Knoll and Ruska in 1932 led to the discovery of even smaller, never-before-seen cell organelles and paved the way for a colossal revolution in the field of biology.
In order to better understand the significance of a cell, let us now take a look at the famous cell theory.
Cell theory
In 1838, a German botanist named Matthias Schleiden closely studied a large number of plants and noticed that all of them had one thing in common – namely, all of them were composed of different types of cells, which were further organized to form tissues. About a year later, a British zoologist named Theodore Schwann (1939) studied various animal cells and discovered that cells possess a thin outer layer that is known today as the plasma membrane.
After conducting several meticulous studies on planet tissues, Schwann also concluded that the presence of cell wall is a unique feature of plant cells. Based on these observations, he proposed the cell hypothesis that states that the bodies of animals and plants are composed of cells and products of cells. Together, Schleiden and Schwann formulated the world-famous cell theory that revolutionized the field of biology.
However, the cell theory was unable to explain how new cells were formed. In 1855, Rudolf Virchow stepped in to rectify this flaw and first explained that new cells are formed from pre-existing cells (“Omnis cellula-e cellula”) by the process of cell division. He proceeded to modify Schleiden and Schwann’s hypothesis to give a final shape to the cell theory.
Today, we understand the cell theory according to the following postulates:
- All living organisms are composed of cells and products of cells.
- All cells arise from pre-existing cells.
A brief outline of the living cell
Have you ever observed the cells in an onion peel or a buccal smear containing human cheek cells under a microscope during your school days? If yes, then let’s discuss a bit about their structure. In typical plant cells such as onion cells, a clear and rigid cell wall serves as the outer boundary and the cell membrane lies just within its confines.
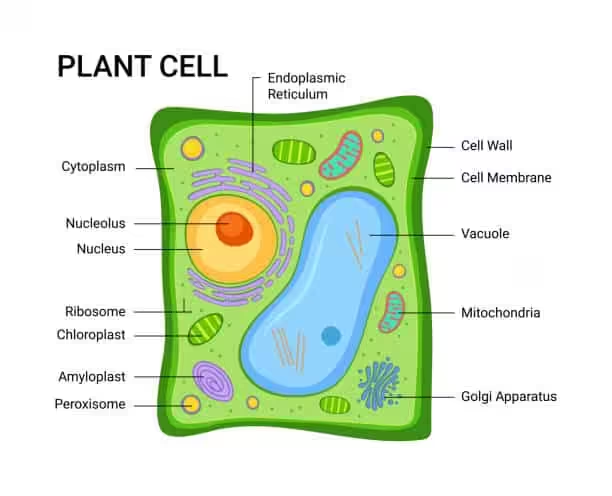
On the other hand, the cell membrane is the delimiting structure of typical animal cells such as human cheek cells. A dense membrane-bound structure known as the nucleus lies within each cell. The nucleus, in turn, contains chromosomes that harbor the genetic material of the cell – namely, the DNA.
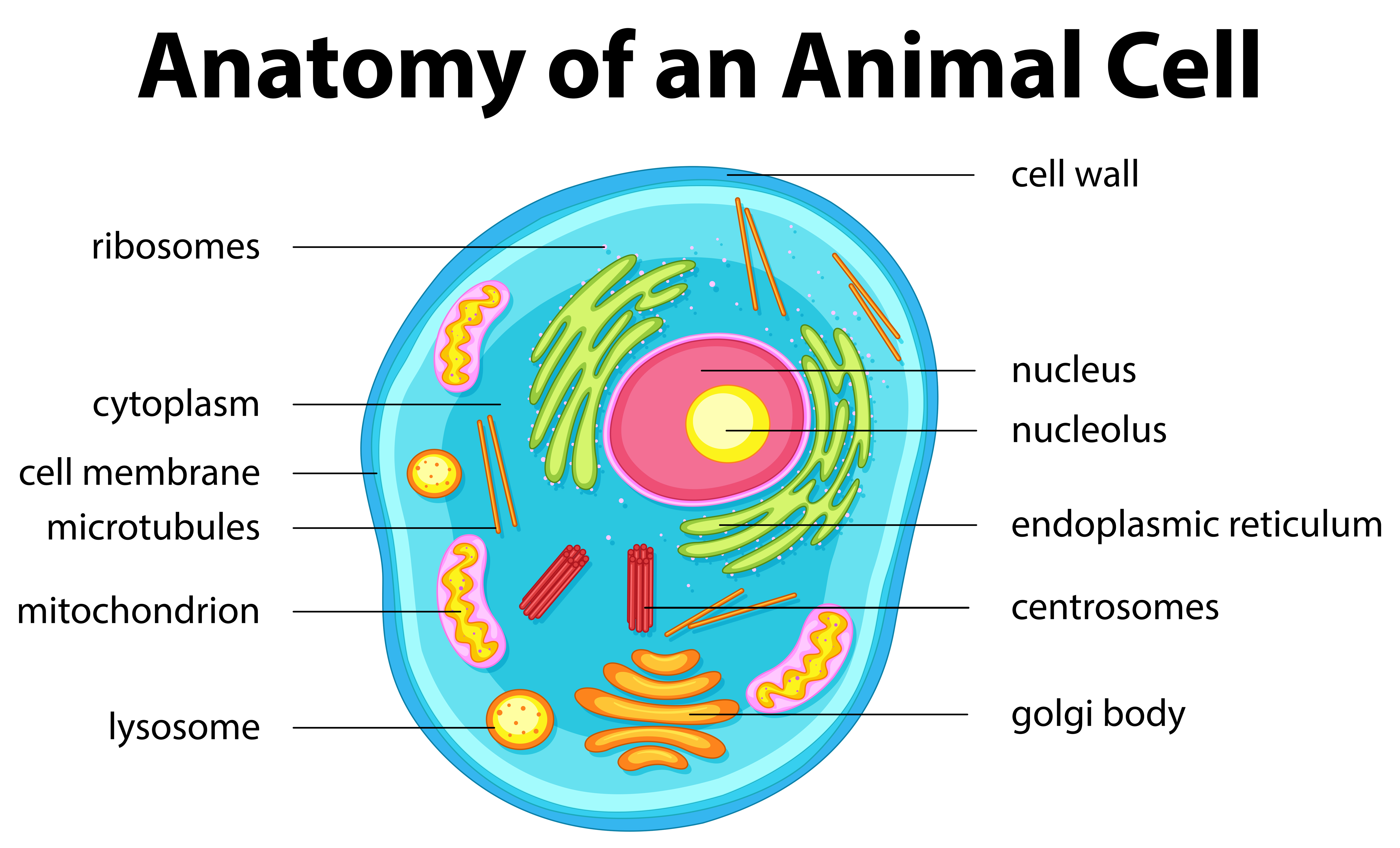
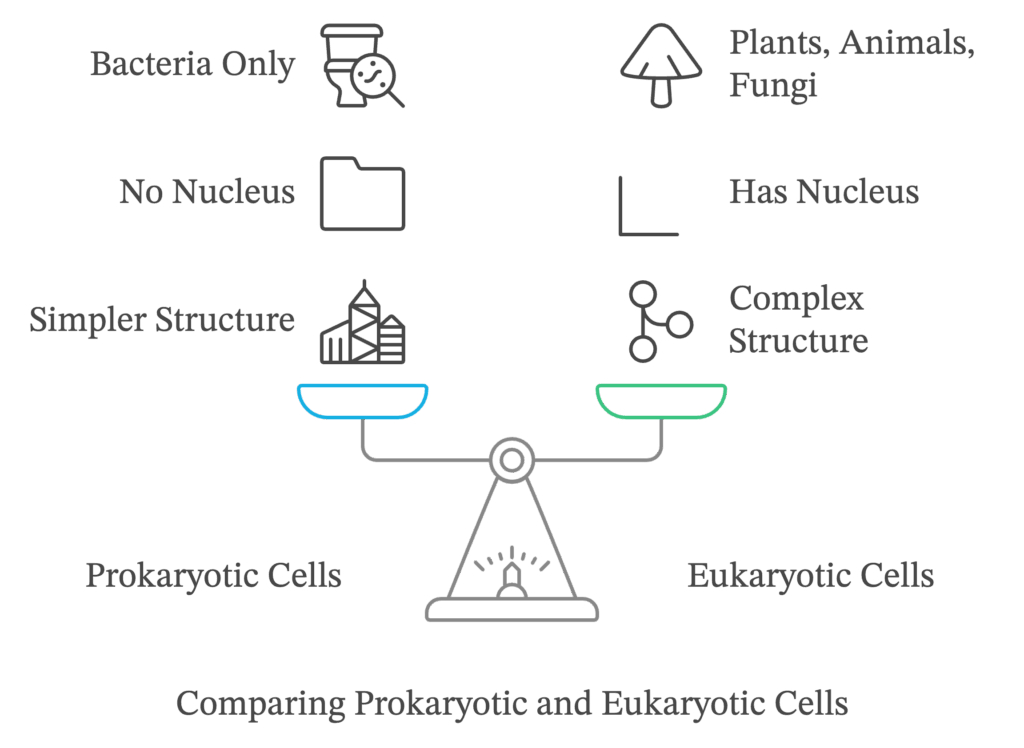
Cells whose nuclei are bound by a distinct nuclear membrane are known as eukaryotic cells, whereas those that lack a membrane-bound nucleus are known as prokaryotic cells. The bulk of the volume of the cell, whether it is prokaryotic or eukaryotic, is occupied by a semifluid matrix known as the cytoplasm. It is the main arena of cellular activities in both plant and animal cells.
Apart from the nucleus, eukaryotic cells also possess other distinct membrane-bound structures known as cell organelles. These include the Golgi complex, endoplasmic reticulum (ER), mitochondria, lysosomes, vacuoles, and microbodies. Prokaryotic cells are notable for lacking membrane-bound organelles.
However, both prokaryotic and eukaryotic cells possess certain non-membrane-bound organelles known as ribosomes. Within the cell, they are found not only in the cytoplasm but also inside two other organelles – chloroplasts (in plants), mitochondria, and on the surface of the rough endoplasmic reticulum (RER). Animal cells possess another unique non-membrane-bound organelle called the centriole which helps in carrying out cell division.
When it comes to size, shape and functions, cells are extremely diverse. For example, the smallest known cell – Mycoplasma – measures only 0.3μm in length, whereas bacteria could measure somewhere between 3 and 5μm. The largest isolated single cell as of now is the egg of an ostrich. Speaking of multicellular organisms, the diameter of human red blood cells is around 7.0μm. Similarly, nerve cells are among the longest cells in existence.
Cells are highly diverse in terms of shape as well. Depending on the function they perform, they could be polygonal, cuboid, columnar, thread-like, disc-like, or even irregular in shape.
Structure of a prokaryotic cell
In the living world, prokaryotic cells are represented by bacteria, cyanobacteria or blue-green algae, Mycoplasma, and pleuro pneumonia-like organisms (PPLO). Compared to eukaryotic cells, they are usually smaller in size but tend to multiply much faster. Though most of them possess a similar basic cellular organization, they could vary greatly in terms of size and shape. The four basic shapes of bacteria are:
- Bacillus (rod-like)
- Coccus (spherical)
- Vibrio (comma-shaped)
- Spirillum (spiral)
We get to see a wide variety of shapes and functions among prokaryotic organisms, but the organization of their cells is fundamentally similar. They usually have a cell wall surrounding the cell membrane, and the cytoplasm serves as the fluid matrix filling the cell. They lack a well-defined nucleus; the genetic material is essentially naked and not enveloped by a nuclear membrane.
A single chromosome or circular DNA molecule serves as the genomic DNA of bacteria. Apart from that, many of them possess small circular DNA molecules outside the genomic DNA, known as plasmids. By virtue of these DNA molecules, bacteria develop certain special phenotypic characters such as resistance to antibiotics. Plasmids are also used by scientists to monitor bacterial transformation with foreign DNA in genetic engineering procedures.
Keep in mind the prokaryotic cells don’t possess any organelles like the ones in eukaryotic cells, except for ribosomes.
Bacterial cell envelope and its modifications
Prokaryotic cells, especially bacterial cells, tend to have a chemically complex cell envelope that consists of three different tightly bound layers:
- The outermost glycocalyx
- The cell wall
- The plasma membrane
The glycocalyx is the outermost layer that features a coating of mucus or polysaccharides that protects the cells and also aids in adhesion. It varies quite a lot in terms of thickness and chemical composition in different bacteria. In some of them, it is present in the form of a loose sheath known as the slime layer that prevents the cell from losing water and valuable nutrients.
In others, it occurs as a thick and tough covering known as the capsule. The slime layer and capsule are both composed of polysaccharides and occasionally proteins. The latter confers a sticky and gummy character to the cell and also helps the bacterium evade the host’s immune system.
The cell wall determines the shape of the cell and provides firm structural support to prevent the bacterium from collapsing or bursting. It is rigid because of the presence of a unique macromolecule known as peptidoglycan or mucopeptide. Numerous antibiotics such as penicillin inhibit the cross-linking of peptidoglycan strands and induce lysis of the bacterial cell, thus helping in the treatment of the disease.
Gram staining
Developed by histologist Hans Christian Gram in 1884, Gram staining is a special technique used to stain and classify bacteria into two different groups – Gram-positive and Gram-negative bacteria. On staining, Gram-positive bacteria turn purple whereas Gram-negative bacteria stain pink because of the fundamental differences in the structure of their cell walls. About 60% to 90% of the
The cell wall of Gram-positive bacteria is made up of peptidoglycan and teichoic acid, whereas only 10% to 20% of the Gram-negative bacterial cell wall is composed of peptidoglycan, arranged in two or three layers. The Gram-negative cell wall is also surrounded by an outer membrane composed of phospholipids, lipoproteins, lipopolysaccharide, and various other proteins.
For carrying out the Gram-staining procedure, we require the following:
- A compound light microscope
- A slide with bacterial smear
- Crystal violet or gentian violet (basic dye)
- 0.5% Gram’s iodine (mordant)
- 95% ethyl alcohol (decolorising agent)
- 1% safranin or dilute carbol fuchsin (counter stain)
The Gram-staining is then carried out as shown below:
- Air-dry the bacterial smear and cover it with crystal violet or gentian violet. Allow it to stand for one minute.
- Rinse the smear gently under mildly flowing tap water.
- Cover the smear with 0.5% Gram’s iodine and allow it to stand for one minute. It will result in the formation of a Crystal Violet Iodine (CVI) complex.
- Once again, rinse the smear gently under tap water.
- Proceed to decolorise the smear with 95% ethyl alcohol. Hold the slide with your hand on one corner in a slanted position, and add ethanol dropwise until you can observe a faint color coming out of the smear.
- Gently rinse the smear again under mildly flowing tap water.
- Cover the smear with dilute carbol fuchsin for 30 seconds to one minute.
- Rinse the smear gently under tap water, and air dry it.
- Put the slide under the microscope and first observe it under the low power (10x) objective, and then under the oil immersion (100x) objective.
Bacteria that retain the violet stain after decolorizing are Gram-positive. The others that don’t do so and take up the counterstain appear pink in color, and are known as Gram-negative bacteria.
The basis of the Gram reaction is the permeability of the bacterial cell wall and cytoplasmic membrane, to the dye-iodine complex. In Gram-positive bacteria, theCVI complex combines to form a larger molecule that precipitates inside the cell. Moreover, ethyl alcohol causes dehydration of the multi-layered peptidoglycan in the bacterial cell wall.
As a result, the space between the molecules decreases and causes the cell wall to trap the CVI complex within the cell. Thus, Gram-positive bacteria do not get decolorised and retain the primary dye, appearing violet. They also have more acidic protoplasm and thus bind to the basic dye more firmly.
Differences between Gram-positive and Gram-negative bacteria
The differences between Gram-positive and Gram-negative bacteria are of great medical and pharmacological significance. Thus, it is important to take a detailed look at them.
| Gram-positive bacteria | Gram-negative bacteria |
|---|---|
| The bacteria retain their purple color even after decolorizing with ethyl alcohol. | The bacteria don’t retain the primary stain when decolorized with ethyl alcohol. |
| The cell wall is 20-80 nm thick and smooth | The cell wall is 8-12 nm thick and wavy. It only comes in contact with the plasma membrane at a few loci. |
| Outer membrane is absent. | Outer membrane is present. |
| Murein content is 70-80%. | Murein content is 10-20%. |
| Teichoic acid is present in the cell wall. | The cell wall lacks teichoic acid. |
| Basal body of the flagellum contains two rings – S and M. | Basal body of the flagellum contains four rings – L, P, S, and M. |
| Only a handful of pathogenic bacteria are Gram-positive in nature. | Most known pathogenic bacteria are Gram-negative in nature. |
| Examples: Bacillus, Lactobacillus, Clostridium, Staphylococcus, Streptococcus, Corynebacterium, Leuconostoc | Examples: E.coli, Acetobacter, Vibrio, Salmonella, Shigella, Azotobacter, Xanthomonas, Shigella |
Plasma membrane and cell appendages
The plasma membrane is selectively permeable in nature and readily interacts with the world outside the cell. It is structurally similar to the plasma membrane present in eukaryotic cells, which we will discuss further. However, we must take note of a unique membranous structure known as the mesosome here. It is formed by extensions of the plasma membrane into the cell in the form of tubules, lamellae, and vesicles.
Mesosomes aid in the formation of the cell wall, DNA replication and distribution to the daughter cells after cell division. They also play a role in secretory processes, respiration, in increasing the surface area of the plasma membrane and enzymatic content. Certain prokaryotes such as cyanobacteria also possess other membranous, pigment-containing extensions into the cytoplasm known as chromatophores.
Bacterial cells can be either motile or non-motile in nature. Motile bacterial cells possess thin, filamentous extensions protruding from their cell wall known as flagella. Bacteria are highly diverse when it comes to the number and arrangement of flagella. An individual flagellum is composed of three distinct parts –
- Filaments
- Hook
- Basal body
The filament is the longest portion of the flagellum and extends from the cell surface to the outside. It is a rigid, hollow, and cylindrical structure composed of a unique protein known as flagellin. The basal body is a rod-like structure consisting of rings.
Apart from flagella, bacteria also possess other surface structures known as pili and fimbriae that don’t play a role in the motility of the cell. The pili are elongated tubular structures composed of a unique protein known as pilin.
As of now, scientists have found true pili only in Gram-negative bacteria, where they play an important role in bacterial conjugation. Because the formation of pili is usually controlled and specific for every cell type, conjugation only takes place between compatible bacterial cells.
On the other hand, fimbriae are small bristle-like fibers sprouting out of the bacterial cell. They resemble slender tubes composed of helically arranged protein subunits that are 3 to 10 nm in diameter. In some bacteria, they have been observed to help in attaching the bacterial cell to rocks in flowing streams and to host tissues.
Ribosomes and inclusion bodies
Ribosomes are associated with the plasma membrane in prokaryotic cells. They are around 15 nm × 20 nm in size and consist of two different subunits – the 50S and 30S units which, when joined together, form 70S prokaryotic ribosomes. Ribosomes are the sites of protein synthesis in the cell. Proteins synthesized by ribosomes in the cytoplasm remain within the cell, but ribosomes on the plasma membrane produce proteins that are subsequently transported out. When many ribosomes attach to a single mRNA, they form a chain-like structure known as a polyribosome or polysome. Within the ribosomes of a polysome, the mRNA is translated into proteins.
Inclusion bodies are essentially aggregations of reserve material in prokaryotic cells stored in the cytoplasm. Most of them are not membrane-bound and lie free in the cytoplasm. However, a few types of inclusion bodies may be surrounded by a single layer non-unit membrane which is around 2-4 nm thick. Examples of inclusion bodies include glycogen granules, cyanophycean granules, and phosphate granules. Blue-green, purple and green photosynthetic bacteria have gas vacuoles as well.
Structure of a eukaryotic cell
Eukaryotic cells possess an organized nucleus bound by a nuclear envelope. Because of the presence of membrane-bound organelles, we see extensive compartmentalization of cytoplasm in these cells. Moreover, they feature a wide variety of complex cytoskeletal structures and appendages for locomotion. Unlike prokaryotic cells, their genetic material is organized into distinct chromosomes. Examples of eukaryotic cells include all the protists, animals, and fungi. However, not all eukaryotic cells are identical to each other.
The cell envelope in eukaryotes consists of both a cell wall and a plasma membrane in plants but is formed of only the plasma membrane in animal cells. Also, plant cells possess plastids and a large central vacuole that are absent in animal cells. On the other hand, animal cells have unique structures known as centrioles which are absent in nearly all plant cells. Centrioles play an important role in cell division.
Differences between prokaryotic and eukaryotic cells: a comparison
| Prokaryotic cells | Eukaryotic cells |
|---|---|
| Usually smaller in size. | Comparatively larger in size. |
| Have a single envelope system. | Have a double/two envelope system. |
| Poorly defined nuclear region because of the absence of a nuclear envelope. Genetic material is in direct contact with the cytoplasm, | The nucleus is distinct and bounded by a nuclear membrane. The nuclear material is not in direct contact with the cytoplasm. |
| Nucleolus is absent. | Nucleolus is present. |
| Membrane-bound organelles are absent. | Membrane-bound organelles such as mitochondria, Golgi apparatus, endoplasmic reticulum, lysosomes, and plastids (in plants) are present. |
| Protein synthesis takes place only in the cytoplasm. | Protein synthesis takes place in the cytoplasm, mitochondria, and plastids. |
| Ribosomes are of 70S type. | Ribosomes are of 80S type. 70S type of ribosomes is found in mitochondria and plastids. |
| Centrioles are absent. | Centrioles are present in animal cells. |
| True sap vacuoles are generally absent, and gas vacuoles may be found in their place. | True sap vacuoles are commonly seen in plant cells. |
| Plasmids may be present. | Plasmids are absent. |
| Thylakoids, if present, lie freely in the cytoplasm. | If present, thylakoids are stacked inside the chloroplast to form a granum. |
| Possess a single envelope system. | Possess a double/two envelope system. |
Let us now take an individual look at the various cell organelles and understand their structure and functions in detail.
Cell membrane or plasma membrane
The cell membrane or plasma membrane is responsible for the relationship of a cell with the outside world. Thus, it is absolutely necessary for the survival of the cell. Scientists managed to devise a detailed structure of the plasma membrane only after the invention of the electron microscope by Knoll and Ruska in the 1950s. At the same time, chemical analyses of the cell membrane helped them deduce the possible model of the membrane.
From these studies, they understood that the cell membrane is largely composed of lipids and proteins. The most important lipids in the membrane are phospholipids that are arranged in a bilayer. The lipids are also arranged on the inner side of the membrane with their polar heads towards the outer side and their hydrophobic tails towards the inside.
This arrangement protects the nonpolar tail of saturated hydrocarbons from the aqueous environment. Apart from phospholipids, the plasma membrane also contains cholesterol. Biochemical studies later revealed the presence of proteins and carbohydrates in the membrane as well. Depending on the cell type, the ratio of protein and lipid can vary noticeably. For example, the plasma membrane of human erythrocytes (red blood cells) consists of about 52% protein and 40% lipids.
Membrane proteins are classified as integral and peripheral depending on the ease of their extraction from the membrane. Peripheral proteins, which lie on the outer surface of the membrane, can be extracted quite easily. On the other hand, integral proteins are buried partially or fully within the membrane and are much more difficult to remove.
In 1972, Singer and Nicholson proposed a much-improved model of the structure of the cell membrane, widely accepted as the world-famous fluid mosaic model. According to this model, the quasi-fluid nature of the lipid component enables the lateral movement of proteins within the overall bilayer. This ability of the proteins to move inside the membrane is measured as its fluidity.
The fluid nature of the cell membrane plays an important role in functions such as the formation of intercellular proteins, cell division, cell growth, endocytosis, secretion, and so on. The transport of molecules across the membrane is one of its most crucial functions. It is selectively permeable to some molecules present on either side of it.
Transport of substances across the plasma membrane
The passage of substances across the plasma membranes takes place by various methods such as passive transport and active transport. Let us take a closer look at these processes:
Passive transport – Several molecules move across the membrane without any expenditure of energy; this process is known as passive transport. Natural solutes move across the membrane along the concentration gradient (i.e., from higher concentration to lower concentration) by the process of simple diffusion.
The diffusion of water across the membrane along the concentration gradient takes place by a process known as osmosis. Since polar molecules cannot pass through the non-polar lipid bilayer, they require a special carrier protein to facilitate their transport across the membrane.
Active transport – It is defined as an uphill movement of substances across the plasma membrane. Here, solute particles move against the concentration gradient – i.e., from their lower concentration to higher concentration. This is the process that requires the utilization of energy, which is obtained from ATP. A good example of active transport is the Na+/K+ ATPase pump in animal cells.
Another kind of active transport is bulk transport, which involves the transport of large amounts of substances such as lipid droplets and solid food particles across the plasma membrane with the expenditure of energy. It is mainly of two types – endocytosis and exocytosis.
Endocytosis involves the intake of substances in the form of carrier vesicles formed by the invagination of tiny regions of the cell membrane. It is of two types – pinocytosis and phagocytosis.
- Pinocytosis – It is also known as “cell drinking”, because it allows high molecular weight fluid substances (fats, proteins, amino acids, lipoproteins, insulin, and so on) to enter the cytoplasm in the form of globules of fluid. These globules, known as pinosomes, are pinched off from the cell membrane inside the cytoplasm in the form of tiny pinocytic vesicles.
- Phagocytosis – Also known as the cell eating process, it is the bulk intake of large solid particles by the cell with the help of the cell membrane. The ingested particle is pinched off from the membrane inside the cytoplasm in the form of a food vacuole or phagocytic vesicle that is later digested by lysosomes. Phagocytosis is commonly seen in all protozoans and in special cells of metazoans such as leucocytes or white blood cells, the reticular cells of the spleen, and so on.
Exocytosis is the process of throwing out the undigested waste products or secretory products from the cell’s cytoplasm with the help of the cell membrane. It is also known as cell vomiting or exphagy.
Cell wall
Like we discussed earlier, the cell wall is a non-living and rigid structure that forms an outer covering for the cell membrane of plants and fungi. It gives a definite shape to the cell and protects it from infection and mechanical trauma. It also plays an important role in cell-to-cell interaction and serves as a barrier to undesirable and potentially harmful macromolecules.
The composition of the cell wall also varies depending on the group we’re dealing with, as shown below:
- Fungal cell wall – The cell wall of fungi is usually composed of chitin, a polymer of N-acetyl glucosamine (NAG).
- Algal cell wall – Algae possess a cell wall composed of cellulose, galactans, mannans, and minerals like calcium carbonate.
- Plant cell wall – The cell wall in other plants is composed of cellulose, hemicellulose, pectins, and proteins.
The cell wall of plants consists of two different regions – the primary wall and the secondary wall.
- Primary well – Young plant cells have a thin and elastic primary wall that is capable of expansion in growing cells. It grows by the addition of more wall material within the already existing one. This type of wall is found in meristematic and parenchymatous cells.
- Secondary wall – The capacity of the primary wall to grow gradually decreases as the cell matures. Thereafter, a secondary wall is formed by the addition of more layers of substances like lignin and suberin internal to the primary wall (towards the plasma membrane). This results in the thickening of the cell wall and is generally seen in cells that form the harder woody parts of plants.
Also, adjacent plant cells in a plant tissue are held together by a thick, amorphous, and sticky layer of cementing material known as the middle lamella. It is primarily composed of calcium and magnesium pectate. In ripening fruits, these pectate compounds solubilize to form a jelly-like mass that results in the fruits becoming soft.
The cell wall and middle lamella could be traversed by the plasmodesmata – a structure that connects the cytoplasm of neighboring plant cells.
Functions of the cell wall
The cell wall carries out many important functions, which have been shown below:
- It maintains the shape of the cell.
- It protects the cell from mechanical trauma.
- It allows various substances to pass in and out of the cell.
- It serves as a barrier against pathogens like bacteria, viruses, fungi, and so on.
- It facilitates cell-to-cell interaction and keeps away undesirable macromolecules.
The thickness of the cell wall is not uniform throughout, because the cell wall is not laid down at certain places. Such relatively thinner areas are known as pits. Pits of adjacent cells usually lie opposite to each other and form pit pairs.
Pits are of two different types –
- Simple pits – In these, the pit cavity has a uniform diameter.
- Bordered pits – They have a flask-shaped pit cavity like in tracheids.
Plasmodesmata is the sole living component of the otherwise dead cell wall. A large number of plasmodesmata or cytoplasmic strands are present in pits, though which the cytoplasm of one cell is in contact with that of another cell. They’re lined by the plasma membrane and contain a fine tubule known as the desmotubule. The endoplasmic reticulum plays an important role in the origin of plasmodesmata as well. Together, they lead to the formation of the symplastic system.
The endomembrane system
There are many membrane-bound organelles present inside a eukaryotic cell. Each of them has a distinct structure and carries out a specific set of functions. However, some of these organelles work in a coordinated manner and lead to the formation of an endomembrane system. This system comprises of the following organelles:
- The endoplasmic reticulum
- The Golgi complex
- Lysosomes
- Vacuoles
Since the functions of chloroplasts, mitochondria, and peroxisomes aren’t coordinated with those of the organelles mentioned above, they are not considered to be a part of the endomembrane system.
We’ll now discuss every component of the endomembrane system in detail below.
The endoplasmic reticulum
After performing electron microscopic studies of eukaryotic cells, scientists noticed the presence of a network or reticulum of tiny tubular structures scattered in the cytoplasm. This network, known as the endoplasmic reticulum or ER, divides the intracellular space into two distinct compartments –
- Luminal compartment – This is the internal space enclosed by the ER membrane.
- Extraluminal compartment – It is the space outside the ER in the cytoplasm.
The ultrastructure of the endoplasmic reticulum is composed of three types of structures – cisternae, tubules, and vesicles. We’ll discuss each of them in detail below.
- Cisternae – These are long, parallel, flattened, interconnected, sac-like structures found in cells that are actively involved in protein synthesis. They are generally found in cells that have synthetic roles, such as cells of the pancreas and brain. Also, they tend to be associated with the large 60S subunit of ribosomes.
- Tubules – They are either branched or unbranched structures that form the reticular system alongside the vesicles and cisternae. They’re not associated with ribosomes, and are commonly found in cells involved in lipid and sterol synthesis.
- Vesicles – These are oval, membrane-bound vacuolar structures. Like tubules, they are not associated with ribosomes. They are found abundantly in the cells of the pancreas. Also, they are notable for being the only ER structures to be found in spermatocytes.
Depending on the presence or absence of ribosomes on the surface of the endoplasmic reticulum, it can be of two kinds:
- Rough/granular
- Smooth/agranular
(a) Rough endoplasmic reticulum (RER)
It is the endoplasmic reticulum that bears ribosomes on its surface and has a rough, granular appearance under the electron microscope as a result. They’re extensive and continuous with the outer nuclear membrane. The RER has two different kinds of glycoproteins – ribophorin-I and ribophorin-II for the attachment of the 60S subunit of 80S ribosomes.
Since ribosomes are the sites of protein synthesis, the RER is found in cells that are actively involved in protein synthesis and secretion. It also gives rise to the smooth endoplasmic reticulum (SER) and provides precursors of certain enzymes for the formation of lysosomes in the Golgi complex.
(b) Smooth endoplasmic reticulum (SER)
The SER is the endoplasmic reticulum that is free of ribosomes. It appears as a network of smooth, tubular structures when observed under the electron microscope. Muscle cells possess large amounts of a special kind of SER known as the sarcoplasmic reticulum.
The SER is mainly involved in the synthesis of lipids and steroids. It also helps in the detoxification of various drugs and passing synthetic products of the RER onto the Golgi complex. The sarcoplasmic reticulum is associated with muscle contraction by the release and uptake of Ca+2 ions.
The Golgi apparatus
The Golgi apparatus or Golgi complex was first observed by Camillo Golgi in 1898. He described it as a network of densely stained, reticular structures present near the nucleus of the cell. They were subsequently named Golgi bodies after their discoverer.
The Golgi apparatus is found in all eukaryotic cells except sperm cells of bryophytes and pteridophytes, mature sieve tubes of plants, and mature RBCs of mammals. It is absent in prokaryotic cells. Also, many fungi and ciliated protozoans lack well-formed Golgi bodies.
In plants, the Golgi complex is composed of unconnected units and is known as dictyosomes. It consists of four different parts – cisternae, tubules, vesicles, and Golgian vacuoles. Let us study each of them in detail.
(a) Cisternae
Cisternae give rise to an extensive network and are arranged in a concentric pattern near the nucleus. They’re flattened sac-like structures that are stacked upon each other. Generally, there are 4-8 cisternae present in every stack. They closely resemble the SER in terms of appearance. In fungi, Golgi bodies are unicisternal.
The number, shape, and size of cisternae could vary in different cells. However, they have a similar organization in every particular type of cells. Their size ranges from 0.5μm to 1.0μm in diameter. They’re arranged concentrically near the nucleus with a distinct convex cis or the forming face and a concave trans or the maturing face.
Although the cis and the trans faces of the organelles are completely different, they are interconnected.
(b) Tubules
Tubules are small, flat, interconnecting structures that arise from the periphery of cisternae due to fenestrations.
(c) Vesicles
Vesicles are large, rounded sacs located at the edges of cisternae in the form of clusters. They are pinched off from the tubules and are of two types:
- Smooth vesicles – As their name suggests, they are secretory vesicles with smooth surfaces that contain secretory granules.
- Coated vesicles – They’re spherical protuberances with rough surfaces arising from the tubules of cisternae.
(d) Golgian vacuoles
These are large, spherical vacuoles that are produced at the maturing face and are filled with some amorphous or granular substances. Some of them also function as lysosomes.
Functions of the Golgi apparatus
- The most important function of the Golgi apparatus is to process, package, and transport the substances for secretion. The packaged material is delivered either to the intracellular targets (within the cell) or secreted to extracellular targets (outside the cell).
The material to be secreted moves from the ER to the Golgi complex in the form of transitional vesicles, which subsequently fuse with the cis face and move towards the maturing face of the Golgi complex. Thus, the Golgi apparatus is in close association with the ER both structurally and functionally.
- Various proteins synthesized by ribosomes present on the ER are transferred to the Golgi complex. They’re then modified in the cisternae of the Golgi apparatus before being released from the trans face.
- It is the most important site of the formation of glycoproteins (by the glycosylation of proteins) and glycolipids (by the glycosylation of lipids).
- The acrosome of the sperm is essentially a modified Golgi apparatus.
- Root cap cells possess abundant Golgi bodies that secrete mucilage for the lubrication of the root tip.
- It helps in the formation of the plasma membrane during cytokinesis.
Lysosomes
Lysosomes are spherical, membrane-bound, vesicular structures formed by the process of packaging in the Golgi complex. They’ve been found to be rich in almost all kinds of hydrolytic enzymes (hydrolases – proteases, lipases, carbohydrases) that are optimally active at an acidic pH. They’re capable of digesting proteins, lipids, carbohydrates, and nucleic acids.
H+ ions are pumped into lysosomes to maintain the acidic conditions within them. Lysosomes are notable for displaying polymorphism; on the basis of their morphology, contents, and functions, they are divided into four different forms:
(a) Primary lysosomes – They’re tiny, vesicle-like newly formed structures produced from the trans face of the Golgi complex. They contain inactive enzymes.
(b) Secondary lysosomes – They’re also known as heterophagosomes or digestive vacuoles. They’re formed when phagosomes fuse with pre-existing primary lysosomes. They contain appropriate enzymes to act upon the substances to be digested.
(c) Residual bodies – They are secondary lysosomes that contain undigested materials. Residual bodies move outwards, come in contact with the plasma membrane, and discharge their contents outside through the process of exphagy or exocytosis. In some cells such as muscle cells and liver cells, this process doesn’t take place; instead, residual bodies load the cells and bring about ageing.
The absence of exphagy from residual bodies may result in polynephritis. Various other diseases such as lung fibrosis, gout, and arthritic joints are associated with the malfunctioning of lysosomes. Around 20 genetic diseases in human beings have been known to occur due to the deficiency of certain lysosomal enzymes. Examples of such diseases include Farber’s disease, Niemann-Pick disease, and Hunter’s syndrome.
(d) Autophagic vacuoles – These are formed by the union of several primary lysosomes around old or dead organelles. The lysosomes surround them with a vacuolar membrane and digest them by the process of autodigestion or autolysis. They’re also known as suicidal bags. Autolysis plays an important role in the disappearance of larval organs during metamorphosis, such as the tail in frogs.
Vacuoles
The vacuole is a membrane-bound space in the cytoplasm containing water, excretory products, sap, and other substances not useful for the cell. They’re also known as sap vacuoles. In plant cells, they may occupy up to 90% of the volume of the cell.
They’re bounded by a single semipermeable membrane known as the tonoplast that facilitates the transport of various ions and other substances against their concentration gradients into the vacuole. As a result, their concentration in the vacuole is noticeably higher than that in the cytoplasm.
The different types of vacuoles are:
- Contractile vacuole – It helps in excretion in Amoeba.
- Food vacuoles – They’re formed in many cells, such as protists, after the engulfing of food particles.
- Gas vacuoles – They’re vacuoles without membranes that provide buoyancy in prokaryotes.
Mitochondria
Mitochondria are cylindrical or sausage-shaped, double-membraned organelles that are found scattered throughout the cytoplasm. Since they’re difficult to view under the microscope, they are first stained by a vital stain known as Janus green to make them visible enough for observation.
Mitochondria are quite variable in terms of their shape, size, and number. A typical mitochondrion has a diameter in the range of 0.2μm to 1.0μm (an average of 0.5μm), and a length ranging from 1.0μm to 4.1μm. Their number also varies greatly in different cell types depending on the amount of work done by the cell and its energy needs.
Structure of a mitochondrion
Mitochondria are surrounded by two membranes – an outer and an inner membrane. The outer membrane is smooth and serves as the continuous limiting boundary of the mitochondria. On the other hand, the inner membrane is selectively permeable and forms numerous infoldings known as cristae.
Chemically, the outer membrane is composed of 60% proteins and 40% lipids. The inner membrane has 80% proteins and 20% lipids, and is also rich in cardiolipins. Because of the presence of two membranes, the mitochondrion is partitioned into two different chambers filled with aqueous liquid:
- Outer compartment or intermembrane/pre-mitochondrial space – It is the space between the two mitochondrial membranes.
- Inner compartment or matrix – It is the space bounded by the inner membrane. The cristae are formed towards the matrix and increase the surface area for enhancing enzyme action.
The matrix contains a single, circular, double-stranded DNA molecule with high guanine and cytosine content, 70S ribosomes, some RNA molecules, and all the components required for protein synthesis. It also has necessary enzymes for the tricarboxylic acid (TCA) cycle.
The mitochondria divide by the process of fission. Both the membranes have their own unique enzymes associated with the mitochondrial function. For example, the electron carriers and enzymes required for the formation of ATP are only present in the inner membrane.
The inner face of the inner membrane and the cristae are studded with several knob-like or spherical protuberances known as oxysomes/F1 particles/Fernandez-Moran particles/elementary particles. Every oxysome consists of three parts – the base, stalk, and head piece. The head piece contains the enzyme ATP synthetase for generating ATP via the process of oxidative phosophorylation.
Functions of mitochondria
- Mitochondria serve as the primary sites of aerobic respiration. They’re very much like tiny biochemical factories where food material or respiratory substrates are fully oxidized to carbon dioxide and water. They carry out the oxidation of carbohydrates and proteins and the β-oxidation of fats.
- The energy produced by the mitochondria in the form of ATP is used to perform several energy-dependent processes of the cell such as nerve impulse conduction, muscle contraction, and so on. Mitochondria are known as the power houses of the cell because of their production of ATP.
- In plants, the genes for cytoplasmic male sterility are usually located in the mitochondrial genome.
Plastids
Plastids are commonly found in all plant cells and all euglenoids. By virtue of their large size, they are easily observable under the microscope. Depending on their color and the pigments they have, plastids can be classified into three main types:
(a) Chloroplasts
Chloroplasts are green-colored plastids that contain photosynthetic pigments, chlorophylls, and carotenoids. They actively take part in the synthesis of foodstuffs for the plant. Most of the chloroplasts in green plants are found in the mesophyll cells of the leaves.
They may be 2-4μm thick and have a length of 5-10μm. When it comes to shape, they could be spherical, discoid, oval, lens-shaped, or even ribbon-shaped in some plants. Their number varies from one per cell (in a green alga called Chlamydomonas) to 20-40 per cell in the mesophyll.
Structure of a chloroplast
Similar to mitochondria, chloroplasts are double membrane-bound organelles with an outer and inner membrane. The inner membrane is relatively less permeable to materials than the outer membrane, and thus has a larger amount of proteins, including carrier proteins. The space enclosed by the inner membrane is known as the stroma, which contains numerous organized, membranous, flattened sacs known as thylakoids.
These sacs are arranged in stacks resembling piles of coins known as grana. The thylakoids of different grana are connected by flat, membranous tubules known as the stroma lamellae. The space within the membrane of the thylakoids is known as the lumen. The stroma contains various enzymes needed for the synthesis of carbohydrates and proteins. It also has small, circular, double-stranded DNA molecules and 70S ribosomes.
Keep in mind that these ribosomes are smaller than cytoplasmic (80S) ribosomes.
Functions of the chloroplast
The main function of chloroplasts is photosynthesis – the process of conversion of solar energy into energy-containing carbohydrates. Photosynthesis consists of two phases – the light reaction (performed in thylakoids), and the dark reaction (performed in the stroma). Chloroplasts also store reserve energy in the form of starch granules.
(b) Chromoplasts
Chromoplasts are yellow, orange, or reddish in color due to the presence of carotenoid pigments. The orange color of carrot roots is due to the presence of these plastids. They’re formed from either chloroplasts or leucoplasts. For example, tomatoes and chilies change their color from green to red during ripening due to the transformation of chloroplasts to chromoplasts.
(c) Leucoplasts
Leucoplasts are colorless plastids that are usually found near the nucleus in non-green cells. They tend to be variable in terms of form, size, and stored nutrients. They don’t exist in the form of grana. There are three different types of leucoplasts:
- Amyloplasts – They contain starch and are abundantly found in rice grains, potato tubers, etc.
- Elaioplasts – They store oils and fats, and are found in seeds such as castor.
- Aleuroplasts – They contain proteins. A good example is the aleurone cell layer in maize.
The concept of semi-autonomous organelles
Unlike other cell organelles, mitochondria and chloroplasts are semi-autonomous organelles because of the presence of circular DNA, 70S ribosomes, and various kinds of RNAs (mRNA, tRNA, rRNA). As a result, they’re able to perform protein synthesis and self-duplication. New mitochondria and chloroplasts arise by the division of pre-existing ones (in the case of chloroplasts, from proplastids).
Although they can synthesize some of their own proteins, they are dependent on the cytoplasm for the remaining proteins. They’re believed to be bacterial endosymbionts of the cell, because:
- Their membrane closely resembles that of bacteria (for example, they possess special proteins known as porins).
- They possess their own nucleic acids (circular double-stranded DNA and RNA) and 70S ribosomes.
- They also have the ETS and components necessary for generating ATP.
Ribosomes
In 1953, George Palade first observed ribosomes under the electron microscope as dense particles present in the cytoplasm. They’re non membrane-bound particles that are composed of rRNA and proteins.
Ribosomes consist of two subunits – a large dome-shaped subunit and a smaller cap-shaped subunit that is present above the larger subunit. Both of these subunits remain united with each other by virtue of a certain concentration of Mg+2 ions. When the concentration of Mg+2 ions falls below the critical level, the two subunits get separated.
When the concentration of Mg+2 ions in the matrix rises, two ribosomes get associated with each other to form a dimer. During protein synthesis, numerous ribosomes form a long chain on a common mRNA and lead to the formation of a polysome/ergasome/polyribosome.
Ribosomes are of two kinds – 70S and 80S. Here, “S” is the sedimentation coefficient that indirectly serves as a measure of the size and density of the ribosomes. Prokaryotes have 70S ribosomes whereas eukaryotes have 80S ribosomes. However, chloroplasts and mitochondria have 70S ribosomes as well.
(a) 70S ribosomes are composed of ribonucleoproteins in the ratio of 60:40 (RNA : Protein). They consist of the following subunits:
(i) 30S (smaller subunit) – Has 21 proteins and 16S rRNA.
(ii) 50S (larger subunit) – Consists of 34 protein molecules and 23S and 5S rRNA.
(b) 80S ribosomes are composed of ribonucleoproteins in the ratio of 40:60 (RNA : Protein). They consist of two subunits as shown below:
(i) 40S (smaller subunit) – Consists of 33 protein molecules and a single 18S-rRNA.
(ii) 60S (larger subunit) – Has around 40 protein molecules and three kinds of rRNAs – 28S, 5.8S, and 5S.
Functions of ribosomes
Ribosomes are known as protein factories because they’re the primary sites of protein synthesis in a cell. Free ribosomes synthesize non-secretory proteins, such as enzymatic and structural proteins for use within the cell. On the other hand, ribosomes bound to the ER produce secretory proteins. Freshly synthesized proteins are processed with the help of chaperone proteins.
Cytoskeleton
The cytoskeleton is an elaborate network of minute, filamentous, fibrous, and tubular proteinaceous structures that form the structural framework within the cell. It is found only in eukaryotic cells and is involved in various functions such as motility, mechanical support, and maintenance of the shape of the cell.
The cytoskeleton consists of three different types of structures:
(a) Microtubules – They’re found in the cytoplasmic matrix of all eukaryotic cells and also in cilia, flagella, basal bodies, centrioles, and the mitotic apparatus. They’re hollow, unbranched cylinders usually 25nm in diameter with a hollow core of around 15nm.
The boundary of a microtubule consists of 13 parallel protofilaments, each of which is composed of α and β subunits of tubulin (a non-contractile protein). GTP and Ca+2 ions are required for the assembly and disassembly of microtubules.
Functions of microtubules
- They help in the formation of astral rays and spindles during cell division.
- They form the cytoskeleton of cilia and flagella.
- They help in providing shape, rigidity, motility, and form to the cell.
- They determine the position of the future cell plate.
- They help in the intracellular transport of inorganic ions and nutrients.
- They aid in the anaphasic movement of chromosomes.
(b) Microfilaments – They’re solid, rod-like, unbranched fibrils of indeterminate length. Largely composed of a globular protein called actin, they also possess lesser amounts of a filamentous protein known as myosin. They are often associated with the plasma membrane and form an extensive network in the cytoplasm of the cell.
Functions of microfilaments
- They offer support to the cell membrane.
- In amoeboid cells, they help in the formation of pseudopodia.
- They aid in the formation of cleavage furrow during cell division.
- They represent the contractile system if the cell and play an active role in amoeboid movements and cytoplasmic streaming.
(c) Intermediate filaments – Essentially, they are non-contractile and hollow filaments composed of acidic proteins. They form a basket of sorts around the nucleus and are involved in the formation of scaffolds for chromatin.
Centrioles and centrosome
The centrosome is an organelle that generally contains two cylindrical structures known as centrioles. These are surrounded by a cloud of amorphous pericentriolar material known as the kinoplasm or centrosphere. The two centrioles together are known as the diplosome and are localized in the centrosome at right angles to each other.
The centrioles are not found in higher plant cells but are seen in nearly all eukaryotic cells such as animal cells, algae, and fungi.
Structure of a centriole
A centriole has a whorl of nine evenly placed peripheral fibrils composed of tubulin. There are no fibrils in the centre; thus, the arrangement is known as 9+0. Each fibril is further composed of three subfibres – A, B, and C. Therefore, it is known as a triplet fibril. Adjacent triplet fibrils are connected by C-A proteinaceous linkers.
At the centre of the centriole, there is a rod-shaped proteinaceous mass known as the hub, from which nine proteinaceous strands known as radial spokes arise towards the peripheral triple fibrils. The presence of the peripheral fibrils and radial spokes confers a unique cartwheel appearance to the centriole.
Although the centriole isn’t technically bounded by a membrane, it is surrounded by pericentriolar satellites known as massules (microtubule generators or MTGs).
Functions of the centriole
- Centrioles aid in the formation of spindle bodies that give rise to the spindle apparatus during cell division.
- They help in the formation of basal bodies that give rise to cilia and flagella.
Cilia and flagella
Cilia and flagella are fine, hair-like outgrowths of the cell membrane. Cilia are relatively smaller structures that work like oars and cause the movement of either the cell or the surrounding fluid. On the other hand, flagella are comparatively longer and are responsible for cell movement.
Although they’re found in both prokaryotic and eukaryotic cells, they’re structurally different in both of them. Both the cilia and flagella arise from the centriole-like basal bodies present at their bases that anchor them and control their movements. Let us now study the electron microscopic structure of cilia and flagella in detail.
Structure of cilia and flagella
Cilia and flagella are membrane-bound extensions of the plasmalemma. They’re cylindrical structures and their core, known as the axoneme, has numerous microtubules running parallel to the long axis.
The axoneme is composed of nine microtubule doublets arranged radially along the periphery and a lone pair of microtubules present in the center. This arrangement of microtubules is known as the (9+2) organization. The pair of tubules in the center are connected to each other by a bridge and enclosed by a common sheath known as the central sheath.
Each of the peripheral doublets of microtubules is connected to the central sheath by a radial spoke. Thus, there are a total of nine radial spokes. The peripheral doublets are connected to each other with the help of linkers composed of a special nexin protein.
Differences between cilia and flagella
| Cilia | Flagella |
| The number of cilia per cell is generally quite huge. | Usually, there are 1-4 flagella per cell. |
| They’re shorter in length. | They’re comparatively longer than cilia. |
| They usually occur throughout or on most of the surface of a cell. | They are generally found at one end of the cell. |
| They help in locomotion, circulation, feeding, and so on. | They’re only used for locomotion. |
Nucleus
The nucleus was first discovered by Robert Brown in 1831. Later, Fleming used the term chromatin for the material contained in the nucleus that was stained by basic dyes. The nucleus is a relatively large organelle that is responsible for controlling all the activities of a eukaryotic cell.
Some cells have been known to have more than one nucleus. For example, binucleate cells such as those of Paramecium have two nuclei per cell, while multinucleate cells like in Opalina have many nuclei. Some living cells even lack nuclei at maturity and are known as anucleate, such as sieve tube cells and mammalian red blood cells.
Structure of the nucleus
The nucleus of a non-dividing cell is known as an interphase nucleus. A typical interphase nucleus consists of the nuclear envelope, nucleoplasm, nucleolus, and chromatin fibers.
The nuclear envelope bounds the nucleus on the outside and separates it from the cytoplasm. It is composed of an outer and an inner membrane; the latter is smooth whereas the former is either smooth or rough due to the presence of ribosomes like the RER. This is because the outer membrane is often connected to the ER.
The space between the outer and inner membrane is called the perinuclear space. It is around 10nm to 50nm wide. The nuclear envelope has several complex pores that are formed by the fusion of the outer and inner membranes. These nuclear pores control the passage of materials like proteins, ribosomes, and RNA to outside or inside of the nucleus.
The nucleoplasm is a semifluid, transparent, and colloidal material that fills up the interior of the nucleus. It contains the nucleolus and highly extended, elaborate nucleoprotein fibers known as chromatin.
The nucleolus (plural: “nucleoli”) is a spherical structure found in the nucleoplasm. Since it is a non-membrane bound structure, it is not separated from the rest of the nucleoplasm. Nucleoli are sites for ribosomal RNA (rRNA) synthesis. Thus, they tend to be larger and more numerous in cells actively involved in protein synthesis.
Chromatin
The interphase nucleus contains a loose, diffused and extended network of nucleoprotein fibers known as chromatin. They were thus named after Flemming, in 1879, managed to stain them with certain basic dyes. These fibers condense to form chromosomes.
Chromatin is largely composed of DNA and basic proteins known as histones. It also has RNA and certain non-histone proteins. The histone proteins are involved in the packaging of DNA into compact structures known as chromosomes. We get to observe structured chromosomes in place of the nucleus during different stages of cell division.
Scientists discovered that after staining chromosomes with basic dyes such as fuegel stain or acetocarmine, they were able to observe two different types of regions:
- Heterochromatin region – It is stained darkly during interphase. It is a genetically inactive and highly condensed region that contains tightly packed DNA.
- Euchromatin region – It is stained comparatively lightly during interphase. It is a genetically active region and contains an abundance of loosely packed DNA. This is where transcription takes place.
In higher organisms, the well-organized nucleus harbors a definite number of chromosomes of definite shapes and sizes. For example, every human cell possesses a nearly two meter long thread of DNA distributed among its 23 pairs of chromosomes (a total of 46 chromosomes).
Structure of a chromosome
A chromosome consists of two identical halves known as chromatids that are held together at a certain point known as the centromere. We observe the centromere as a narrow region of the chromosome called the primary constriction. On each side of the centromere, there are disc-like structures known as kinetochores.
The ends of a chromosome are known as telomeres. They seal the ends of chromosomes and prevent them from being shortened or lost. Depending on the position of the centromere, chromosomes may be of four types:
- Metacentric chromosome (median centromere) – The centromere is located at the center and divides the chromosome into two equal arms (isobranchial) during anaphase. These chromosomes appear to be V-shaped.
- Sub-metacentric chromosome (submedian centromere) – In here, the centromere is located slightly away from the center of the chromosome, or nearer to one end of the chromosome. Thus, the chromosome is divided into one longer and one shorter arm and is called heterobrachial. These chromosomes appear to be L-shaped during anaphase.
- Acrocentric chromosome (subterminal centromere) – Here, the centromere is present very close to one end of the chromosome and results in the formation of one very long and one extremely short arm. These chromosomes appear J-shaped during anaphase.
- Telocentric chromosome (terminal centromere) – In this case, the centromere is located at the terminal end of the chromosome. As a result, the chromosome appears to have a single arm. These chromosomes appear I-shaped during anaphase.
Some chromosomes are observed to possess additional constrictions known as non-staining secondary constrictions or NOR (nucleolar organizer) near their ends. The portion of the chromosome beyond the secondary constriction is known as the satellite.
Chromosomes that possess satellites are known as SAT chromosomes and are considered to be marker chromosomes. Scientists have discovered five pairs of SAT chromosomes in human beings.
Giant chromosomes
In certain organisms, chromosomes tend to assume unique structures in some particular tissues. Let’s take a closer look at them:
(a) Lampbrush chromosomes – First described by Ruckert in 1892, these are bivalent diplotene chromosomes joined together at certain points known as chiasmata. They are found in oocyte nuclei of both vertebrates and invertebrates. Their main axis is composed of DNA and each of the bivalents has rows bearing a large number of chromosomes. Lateral projections or loops emerge from many of these chromosomes.
Loops are essentially extended parts of chromosomes that participate in transcription. The hairs present on these loops are actually nascent molecules of RNA. When bound to proteins, they give it a fibrillary and granular appearance. Some of them are stored as combinations mRNA and proteins known as informosomes for use during the development of the embryo.
(b) Salivary gland chromosomes – Also known as polytene chromosomes, they were first observed by E.G Balbiani (1881) in the salivary gland cells of dipteran insects (belonging to the order Diptera) such as Chironomus and Drosophila. By virtue of the presence of these chromosomes, most cytological studies have been performed using Drosophila.
The salivary gland chromosomes in Drosophila have been measured to be up to 2000μm or 2mm! They get their name from the fact that they exhibit polyteny – that is, the number of fibrils or chromonemata increases up to 2000 or more per chromosome. Another characteristic phenomenon called somatic pairing takes place in these chromosomes as well. As a result, their number appears to be half of normal somatic cells.
Microbodies
Scientists have observed numerous single membrane-bound minute vesicles known as microbodies. They’re found in both plant and animal cells and contain various enzymes for catalyzing oxidation reactions other than those of respiration. They include:
- Peroxisomes
- Sphaerosomes
- Glyoxysomes
(a) Peroxisomes – They possess enzymes required for peroxide biosynthesis. They occur in most plants and animals but are found more commonly in photosynthetic cells where they perform photorespiration in conjunction with chloroplasts and mitochondria. They have enzymes for both producing and breaking down peroxide – glycolate oxidase and catalase respectively.
(b) Sphaerosomes – These are unit membrane-bound, refractile, spherical bodies that are produced from the SER. They are involved in the synthesis and storage of fats. Large numbers of sphaerosomes are found in the endosperm cells of oil seeds. They possess hydrolytic enzymes and are believed to be plant lysosomes.
(c) Glyoxysomes – Discovered by Tolbert and Beevers, glyoxysomes are bounded by a single membrane and originate from the ER. They contain enzymes for the glyoxylate cycle where fats are converted into carbohydrates via the process of gluconeogenesis.
They’re commonly found in germinating seeds, especially in germinating fatty seeds such as castor seed and groundnut seed. In these seeds, they turn insoluble lipid food reserves into soluble sugars for the growing plant tip. Since animals don’t have glyoxylate enzymes, they cannot carry out this conversion.